- Buitenwiet test ’26 update #14: code rood
- De voordelen van mulch (bodembedekking) voor wietplanten
- Doe-het-zelf: luchtvochtigheid in de kweektent automatiseren
- The 4th of July: FIFA 420 met DJ Jean & Henk Schulz – Win (5×2) tickets!
- Ongeëvenaarde score politie India met anti-drugs operatie ‘Thunder’
- Wietplanten binnen en buiten beschermen tegen te veel hitte



De creatie van een nieuwe wietsoort: terugkruisen

Alweer het zesde deel van de creatie van een nieuwe wietsoort. Als je deze serie gevolgd hebt dan ken je ondertussen de basis en theorie van het veredelen. Alleen om écht een nieuwe wietsoort te maken, mis je misschien nog de geavanceerde veredel-techniek van het terugkruisen. In dit artikel brengen we je ook daarvan op de hoogte.
Jazeker, in dit artikel doen we een paar geavanceerde veredel-technieken uit de doeken, waarmee je het genotype van wietplanten kunt achterhalen. We zullen je vertellen hoe je F1- en F2-terugkruisingen, F2-hybride kruisingen en F3-terugkruisingen kunt maken. Het doel is hierbij steeds het achterhalen en manipuleren van het genotype van de wietplanten waarmee je werkt. Ken je het genotype van de ouderplanten, dan kun je namelijk voorspellen, welke eigenschappen een bepaalde kruising zal opleveren.
Ken je het genotype van de ouderplanten, dan kun je voorspellen welke eigenschappen een bepaalde kruising zal opleveren
Even herhalen
Dit is zogezegd al het zesde deel van de creatie van een nieuwe wietsoort. We kunnen ons zo voorstellen dat je niet alles helemaal onthouden hebt, dus we nemen nog even door wat we tot nu toe allemaal besproken hebben. Het begint een beetje op een biologieles te lijken. Maar hé, je moet er wat voor over hebben als je een nieuwe wietsoort wil veredelen.
We zijn deze serie begonnen met een aantal basistermen uit de erfelijkheidsleer en hebben daarna het Hardy-Weinberg principe doorgenomen. Een formule waarmee je de frequentie van genotypen voor bepaalde eigenschappen kunt berekenen. In het deel #3 hebben we geleerd hoe we een testkruising kunnen maken om te achterhalen of een eigenschap homozygoot of heterozygoot en dominant dan wel recessief is. In deel #4 en #5 hebben we het tenslotte gehad over Mendels Erwt-experimenten en het maken van genetisch zuivere planten.
Terugkruisen
In het voorbeeld uit deel #5 over het maken van een genetisch zuivere wietplanten, gebruikten we een hypothetische kruising tussen OG Kush en een Neville’s Haze. Beide soorten hadden gewenste eigenschappen voor een fictieve nieuwe soort die we voor het gemak even OG Haze noemden. In het schema hieronder zie je het fictieve resultaat van een eerste F1-kruising van ons voorbeeld.
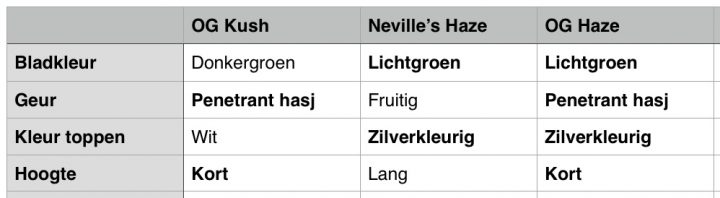
De eerste kruising tussen de Kush en de Haze levert ons zogenaamde F1 hybriden op. Als we er voor het gemak even van uitgaan dat de beide ouders homozygoot zijn voor de eigenschap bladkleur. De OG Kush heeft donkergroene bladeren en de Neville’s Haze heeft lichtgroene bladeren. Stel dat alle F1-hybriden dezelfde lichtgroene bladeren hebben. Dan weten we ook welke eigenschap dominant is, en welke recessief. Bovendien weten we nu van welke plant de dominante dan wel recessieve bladkleur komt, en dat die eigenschappen bovendien homozygoot aanwezig waren in de ouders.

De planten die uit deze kruising voortkomen noemen we F1-hybriden. Omdat S dominant is en s recessief, hebben alle planten de dominante bladkleur. We weten nu ook welke kleur dat is, en van welke ouder die dominante eigenschap kwam. Laten we even stellen dat alle planten lichtgroen van kleur zijn en dat lichtgroen dus dominant is boven donkergroen (S = lichtgroen). De dominante eigenschap voor de bladkleur komt dus van de Neville’s Haze. De recessieve eigenschap voor bladkleur komt van de OG Kush en is donkergroen (s = donkergroen).
Aangezien er in het nageslacht geen variatie zit voor wat betreft de bladkleur, weten we ook dat beide ouders homozygoot waren voor wat betreft de bladkleur (SS en ss). Alle F1-hybriden zijn echter heterozygoot voor wat betreft bladkleur (Ss). Door nu terug te kruizen kunnen we onze genenpoel zo manipuleren dat we 100% lichtgroene bladeren krijgen wanneer we ermee verder veredelen. We kruisen een F1 hybride terug met onze oorspronkelijke Neville’s Haze, zoals je in het schema hieronder ziet.
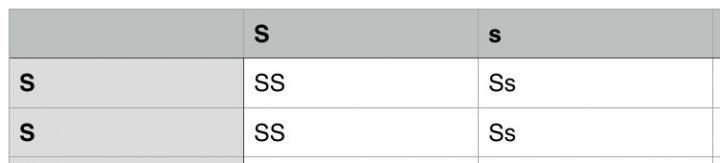
Onze terugkruising resulteert in de zogenoemde F2-terugkruising. Alle planten hebben lichtgroene bladeren, de helft is homozygoot dominant (SS) en de helft is heterozygoot (Ss). Hadden we niet terug gekruist, maar twee heterozygote F1-hybriden met elkaar gekruist, dan hadden we een populatie gehad met 25% homozygoot dominante planten, 50% heterozygote planten en 25 homozygoot recessieve planten. Of met andere woorden: dan had 25% van het nageslacht niet de gewenste lichtgroene bladeren.
Je ziet: terugkruisen is een belangrijke tool uit de gereedschapskist van de zadenveredelaar. Je kunt de frequentie van gewenste fenotypen, er aanzienlijk mee verhogen. De gewone F2 hybride (F1 x F1) levert immers 25% afwijkende fenotypes op met ongewenste donkergroene bladeren. Uit de F2-terugkruising komen alleen maar planten met de gewenste lichtgroene bladeren.
F3-terugkruising
Het voorbeeld hierboven gaat over een simpele terugkruising, maar we kunnen nog verder terugkruisen als we willen. Laten we kijken wat er gebeurt wanneer we onze F2-terugkruising nog eens terugkruisen met de originele ouder. Door stekken van deze planten te nemen kunnen we haar in leven houden terwijl we de F2 kruising maken en het nageslacht daarvan grootbrengen.
Aangezien de F2-terugkruising twee verschillende genotypen opleverden (SS en Ss), zal de helft van onze F3-terugkruising hetzelfde resultaat opleveren als de F2-terugkruising zelf, en de andere helft de gewenste eigenschappen insluiten in planten met die homozygoot dominant zijn voor onze gewenste lichtgroene bladkleur, dat zie je in het schema hieronder.
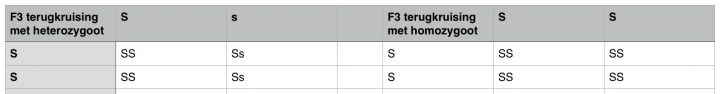
In de tweede F3-terugkruising met de homozygote F2-terugkruising en de originele Neville’s Haze, is al het nageslacht homozygoot dominant (SS) en dus een genetische zuivere plant voor die eigenschap. Welke kruising we verder ook met deze planten nog maken, het resulteert altijd in nageslacht met de gewenste lichtgroene bladeren. De eigenschap is nu zuiver en stabiel.
[Openingsbeeld: annarepp, Shutterstock]
Meer KWEKEN
-

De voordelen van mulch (bodembedekking) voor wietplanten
-

Nice: een van-zaad-naar-oogst docu over Dutch Passions Kerosene Krash
-

Doe-het-zelf: luchtvochtigheid in de kweektent automatiseren
-

Wietplanten binnen en buiten beschermen tegen te veel hitte
-

Living soil (biologische levende aarde) voorbereiden
-

Wanneer is het de moeite waard om een moederplant te kweken?
-

Maak nuttige melkzuur bacteriën voor je wietplanten
-

De grootste kleine fout van beginnende cannabis thuiskwekers
-

Zo toont een wietplant dat hij klaar is om te bloeien (12/12)
-

7 fouten die je moet voorkomen bij het trainen van wietplanten
-

Chitosan: wondermiddel voor wietplanten?
-

Wat ‘het internet’ je over wietzaadjes leert
-

Wietplant hermafrodiet? Herken de eerste signalen & weet wat te doen
-

Zorgt UV-licht nou wel of niet voor meer THC? 70 studies geanalyseerd
-

Hoe veredeling cannabis veranderde & zo maak je een Europees landras!
-

De K.I.S.S. kweekmethode: Keep It Simple …Stupid! ❤
-

6 redenen waarom wietplanten soms nauwelijks groeien
-

Wat is het maximale aantal toppen aan een wietplant?
-

Lichtlekken in je kweektent? Wellicht minder rampzalig dan je denkt
-

Wiet oogsten op basis van de kleur van bloeiharen, hoe betrouwbaar is dat?
-

Studie: langere dagen (18/6) in late bloei levert méér wiet op!
-

Droogtestress voor sterkere wiet: minder water, meer CBD en THC
-

Photobleaching, light (LED) bleach, light burn en lichtstress bij wietplanten
-

Zo ontkiem je ieder wietzaadje! (tenzij het dood is)
-

10 denkfouten die beginnende én ervaren wietkwekers maken



Nu trending op CNNBS
- 1Buitenwiet test ’26 update #14: code rood

- 2Sorry, het is gewoon te heet... hier zijn 512 HIGH & HOT GIRLS

- 3De 'legale cannabis leugen' - waarom iedereen failliet gaat!

- 4Waar zijn Belgen welkom in de coffeeshop? (oktober 2025)
- 5NED vs BEL • 6 beelden uit de oorlog tegen wiet (#98)


- 6Cannabis kweken 2025: compleet kweekplan van week tot week 🌱

- 7De voordelen van mulch (bodembedekking) voor wietplanten

- 8Buiten autoflowers kweken: volledige handleiding met tips & trucs

- 9Wietplanten binnen en buiten beschermen tegen te veel hitte

- 1010 kruiden die je prima kunt mixen met wiet - en de effecten